数学月間の会SGKのURLは,https://sgk2005.org/

数学月間の会SGKのURLは,https://sgk2005.org/
$$\displaystyle \frac{E'}{E}=\displaystyle \frac{1}{1+\alpha (1-cos\theta )}$$
$$\alpha =\displaystyle \frac{E}{m_{0}c^{2 } }$$
クラインー仁科
$$\displaystyle \frac{d\sigma }{d\mit\Omega }=\displaystyle \frac{r_{0 } }{2}\left( \displaystyle \frac{E'}{E} \right) ^{2}\left( \displaystyle \frac{E'}{E}+\displaystyle \frac{E}{E'}-sin^{2}\theta \right) $$
$$E_{e}=E-E'=E\displaystyle \frac{\alpha (1-cos\theta )}{1+\alpha (1-cos\theta )}$$
$$\displaystyle \frac{d\sigma }{dE_{e } }=\displaystyle \frac{d\sigma }{d\mit\Omega }\displaystyle \frac{d\mit\Omega }{d\theta }\displaystyle \frac{d\theta }{dE_{e } }$$
$$ d\mit\Omega =2\pi sin\theta d\theta , \displaystyle \frac{d\mit\Omega }{d\theta }=2\pi sin\theta $$\
$$\displaystyle \frac{dE_{e } }{d\theta }=E\displaystyle \frac{\alpha sin\theta }{\left[ 1+\alpha \left( 1-cos\theta \right) \right] ^{2 } }$$
$$\displaystyle \frac{d\sigma }{dE_{e } }=\displaystyle \frac{d\sigma }{d\mit\Omega }2\pi sin\theta \displaystyle \frac{\left[ 1+\alpha \left( 1-cos\theta \right) \right] ^{2 } }{E\alpha sin\theta }=\displaystyle \frac{\pi r_{0 } }{E\alpha }\left( 1+cos^{2}\theta \right) \left\{ 1+\displaystyle \frac{\alpha ^{2}\left( 1-cos\theta \right) ^{2 } }{\left( 1+cos^{2}\theta \right) \left[ 1+\alpha \left( 1-cos\theta \right) \right] } \right\} $$
1種類の形(2等辺3角形)の赤色と黄色のタイル(赤タイルと黄タイルは互いに鏡像)で作ったタイル張り模様を鑑賞しましょう.1種類の形のタイルで,平面をタイル張りすると,必ず周期的なタイル張りになってしまうと思い込むのは間違っています.確かにFig.4,やFig.5のような周期的なタイリングはすぐ思いつきます.
しかし,Fig.2やFig.3のように非周期なもので,平面をタイル張りするものがあります.Fig.2は中心に回転対称があるタイリング模様で,点群5mの対称性です.Fig.3は,2つの目がある螺旋パターンのタイリングで,水平線は映進面だと思うかもしれませんが,このパターンには周期がありませんから映進操作はできません.
螺旋の目の中間に対称心があります.
さて,ここで万華鏡で作られるタイリング模様Fig.1の登場です.
この万華鏡を生む3枚の鏡は1つの頂点では点群を生成しますが,他の2つの頂点では点群を生成しません.従って平面を赤と黄色の市松模様で埋めることはありません.全体の代数系は,群より緩いもの(特殊な亜群)になってしまいますから非常に複雑です.
対称操作は局所的で,独自の作用域と値域があり興味深いものです.
作用域,値域の制限のために,一つのタイル全体が無傷で写像されるパターン内の位置と,部分が写像される位置があり,このような複雑なタイリング模様ができます.
シュロ縄で柵の竹竿を結びました.庭師は男結びと言う方法で結ぶそうですが,私は簡単にランニング・ノットという方法で結びました.実は,シュロ縄の扱いが大変だったので,一番作業の楽な結び方をして,後でこの結び方の名前を調べたら,ランニング・ノットという方法であることがわかりました.
ランニング・ノット(あるいは,スリップ・ノット)と言われる所以は,竹竿を通してから紐を引っ張って締めると結節ノットが移動して,自然に竹竿の周りの輪が締まるからです.結節になる輪から紐の両端が同じ方向に出ていますから,竹竿を通してから紐の一端を引っ張ると,輪が締り結節になると同時に,他端も同方向に引かれるので,両側から輪を締め,自分自身を締め緩みを防止しする一番シンプルな結び方になります.
ランニング・ノットの結び方で紐の両側を引っ張ると,輪の中に竹竿がなければ手品のように紐は結び目が出来ずに解けてしまいます.比較のために,もやい結びを見てみると,結びの両側を引っ張ると結節ノットは移動せず輪が出来てしまい,竹竿の周りを締める結び方にはなりませんし,竹竿がない状態で,もやい結びの紐の両側を引っ張ると解けずに固定した輪を残して結び目が出来てしまいます.
紐の始まりを竹竿の周りのランニング・ノットから始めて,柵を組んだ竹竿に巻きつけ固定し,紐の最後もランニング・ノットで収めようとするとなかなか難しい.巻いてきたひもが緩まないように締めながら出口の結節になる結び目を作る必要があるからです.
シュロ縄は水に湿らせた方がしなやかでよく締まります.シュロ縄を繰り返ししごいていると,縄に毛玉のような塊や細い箇所ができますから注意しましょう.
今回は,紐の両端をそれぞれ別の場所で固定したので,使いませんでしたが”かます結び”という方法もあります.これは紐の両端を結ぶ結び方です.




■連立線形微分方程式を解く
例えば,次の連立線形微分方程式は,行列を使って表現できます.
$$ \left\{ \begin{array}{@{\,} c @{\, } } \displaystyle \frac{d}{dt}x=y \\[0mm] \displaystyle \frac{d}{dt}y=-x \end{array} \right. $$
$$ \displaystyle \frac{d}{dt}\left[ \begin{array}{@{\,} c @{\, } } x \\[0mm] y \end{array} \right] =\left[ \begin{array}{@{\,} cc @{\, } } 0 & 1 \\[0mm] -1 & 0 \end{array} \right] \left[ \begin{array}{@{\,} c @{\, } } x \\[0mm] y \end{array} \right] \Longleftrightarrow \displaystyle \frac{d}{dt}\vec{x }=M\vec{x } \\ $$
このような微分方程式の解は,初期値を $$\vec{x_{0} } =\vec{x }(t_{0})$$ として,
$$ \vec{x}(t)=\vec{x }_{0}+\displaystyle \int_{t_{0 } }^{t}M\vec{x}(\tau )d\tau $$ となります.
これの計算は,逐次近似で無限の関数列を作れば実行できます.
$$\vec{x_{1 } }(t)=\vec{x}_{0}+M\vec{x_{0 } }(t-t_{0})$$
$$\vec{x_{2 } }(t)=\vec{x}_{0}+\displaystyle \int_{t_{0 } }^{t}M\vec{x_{1 } }(\tau )d\tau =\vec{x}_{0}+M\vec{x_{0 } }(t-t_{0 } )+M^{2}\vec{x_{0 } }\displaystyle \frac{(t-t_{0})^{2 } }{2}$$
$$\vec{x_{n } }(t)=\displaystyle \sum_{0}^{n}M^{n}\vec{x_{0 } } \displaystyle \frac{(t-t_{0})^{n } }{n!}$$
ここで,$$n \longrightarrow \infty $$とすると収束して,次の指数関数の解が得られます.
$$\vec{x }(t)=e^{M\left( t-t_{0} \right) }\vec{x}_{0}$$ ただし,$$e^{Mt} $$の定義は $$e^{Mt}=\displaystyle \sum_{0}^{ \infty }\displaystyle \frac{1}{n!}(Mt)^{n}$$
この解は確かに, $$\displaystyle \frac{d}{dt}e^{Mt}=Me^{Mt}$$を満たします.
■線形化
現実の連立微分方程式は非線形がほとんどです.
平衡点の近傍でテーラー展開し,局所的に方程式を線形化します.
例えば,一般的な反応拡散系の方程式で$$f(u,v),g(u,v)$$は線形とは限りません.
$$ \left\{ \begin{array}{@{\,} c @{\, } } \displaystyle \frac{ \partial u}{ \partial t}=D_{u}\displaystyle \frac{ \partial ^{2}u}{ \partial x^{2 } }+f(u,v) \\[0mm] \displaystyle \frac{ \partial v}{ \partial t}=D_{v}\displaystyle \frac{ \partial ^{2}v}{ \partial x^{2 } }+g(u,v) \end{array} \right. $$
$$u(x,t), v(x,t)$$(それぞれ2種類の物質の濃度)を,平衡点のまわりでテーラー展開し,線形近似します.ただし,平衡点を$$(0,0)$$とする(このようにしても一般性を失わない).
1次の偏微分係数が作る行列(ヤコビアン)$$J$$を定義し,次のように線形化する.
$$ J \equiv \left[ \begin{array}{@{\,} cc @{\, } } \displaystyle \frac{ \partial f}{ \partial u} & \displaystyle \frac{ \partial f}{ \partial v} \\[0mm] \displaystyle \frac{ \partial g}{ \partial u} & \displaystyle \frac{ \partial g}{ \partial v} \end{array} \right] \equiv \left[ \begin{array}{@{\,} cc @{\, } } f_{u} & f_{v} \\[0mm] g_{u} & g_{v} \end{array} \right] $$, $$\left[ \begin{array}{@{\,} c @{\, } } f(u,v) \\[0mm] g(u,v) \end{array} \right] =\left[ \begin{array}{@{\,} cc @{\, } } f_{u} & f_{v} \\[0mm] g_{u} & g_{v} \end{array} \right] \left[ \begin{array}{@{\,} c @{\, } } u \\[0mm] v \end{array} \right] $$
$$f_{u} , f_{v} ,g_{u} ,g_{v}$$は,平衡点$$(0,0)$$での偏微分係数です.
線形化された反応拡散方程式を以下に示します.$$D_{u}, D_{v}$$はそれぞれの拡散係数(常に正).
$$ \left\{ \begin{array}{@{\,} c @{\, } } \displaystyle \frac{ \partial u}{ \partial t}=D_{u}\displaystyle \frac{ \partial ^{2}u}{ \partial x^{2 } }+f_{u}u+f_{v}v \\[0mm] \displaystyle \frac{ \partial v}{ \partial t}=D_{v}\displaystyle \frac{ \partial ^{2}v}{ \partial x^{2 } }+g_{u}u+g_{v}v \end{array} \right. $$
$$u(x,t)=u^{*}e^{\sigma t}\textrm{sin}\alpha x$$, $$v(x,t)=v^{*}e^{\sigma t}\textrm{sin}\alpha x$$ と置くと
$$ \left\{ \begin{array}{@{\,} c @{\, } } \sigma u^{*}=-\alpha ^{2}D_{u}u^{*}+f_{u}u^{*}+f_{v}v^{*} \\[0mm] \sigma v^{*}=-\alpha ^{2}D_{v}v^{*}+g_{u}u^{*}+g_{v}v^{*} \end{array} \right. $$
行列形式で書くと,
$$ \left[ \begin{array}{@{\,} c @{\, } } u^{*} \\[0mm] v^{*} \end{array} \right] =\displaystyle \frac{1}{\sigma }\left[ \begin{array}{@{\,} cc @{\, } } f_{u}-\alpha ^{2}D_{u} & f_{v} \\[0mm] g_{u} & g_{v}-\alpha ^{2}D_{v} \end{array} \right] \left[ \begin{array}{@{\,} c @{\, } } u^{*} \\[0mm] v^{*} \end{array} \right] $$
$$A \equiv \left[ \begin{array}{@{\,} cc @{\, } } f_{u}-\alpha ^{2}D_{u} & f_{v} \\[0mm] g_{u} & g_{v}-\alpha ^{2}D_{v} \end{array} \right] $$
は平衡点$$(0,0)$$におけるヤコビアン.
■解の安定性
$$ \left[ \begin{array}{@{\,} c @{\, } } u^{*} \\[0mm] v^{*} \end{array} \right] =\displaystyle \frac{1}{\sigma }A\left[ \begin{array}{@{\,} c @{\, } } u^{*} \\[0mm] v^{*} \end{array} \right] \Longrightarrow P\left[ \begin{array}{@{\,} c @{\, } } u^{*} \\[0mm] v^{*} \end{array} \right] =\displaystyle \frac{1}{\sigma }PAP^{-1}P\left[ \begin{array}{@{\,} c @{\, } } u^{*} \\[0mm] v^{*} \end{array} \right] $$
$$ PAP^{-1}=\left[ \begin{array}{@{\,} cc @{\, } } \lambda _{1} & 0 \\[0mm] 0 & \lambda _{2} \end{array} \right] $$
が対角化されると,固有値 $$\lambda _{1} , \lambda _{2}$$,固有ベクトルは $$P\left[ \begin{array}{@{\,} c @{\, } }u^{*} \\[0mm]v^{*}\end{array} \right] $$
平衡点 $$(0,0)$$ の解が安定であるためには,すべての固有値が負でなければならない.
$$\lambda _{1}<0,\lambda _{2}<0$$ が必要十分であり,$$\lambda _{1}+\lambda _{2}<0$$では少しゆるい条件になる.
$$A$$の固有値を求めるのは面倒なので,対角化により$$Tr$$は変わらない$$Tr[A]=Tr[PAP^{-1}]$$を利用し,ゆるく評価すると,$$Tr[A]=f_{u}+g_{v}-\alpha ^{2}(D_{u}+D_{v})<0$$
拡散項がない($$D_{u}=D_{v}=0$$)時の安定性から $$f_{u}+g_{v} < 0$$が成立するので,
例えば, $$f_{u}>0, g_{v}<0$$,$$\left| \begin{array}{@{\,} c @{\, } }f_{u}\end{array} \right| <\left| \begin{array}{@{\,} c @{\, } }g_{v}\end{array} \right| $$が得られます.
$$u$$は加速剤,$$v$$は阻害剤として働き,互いの解が安定化することがわかります.
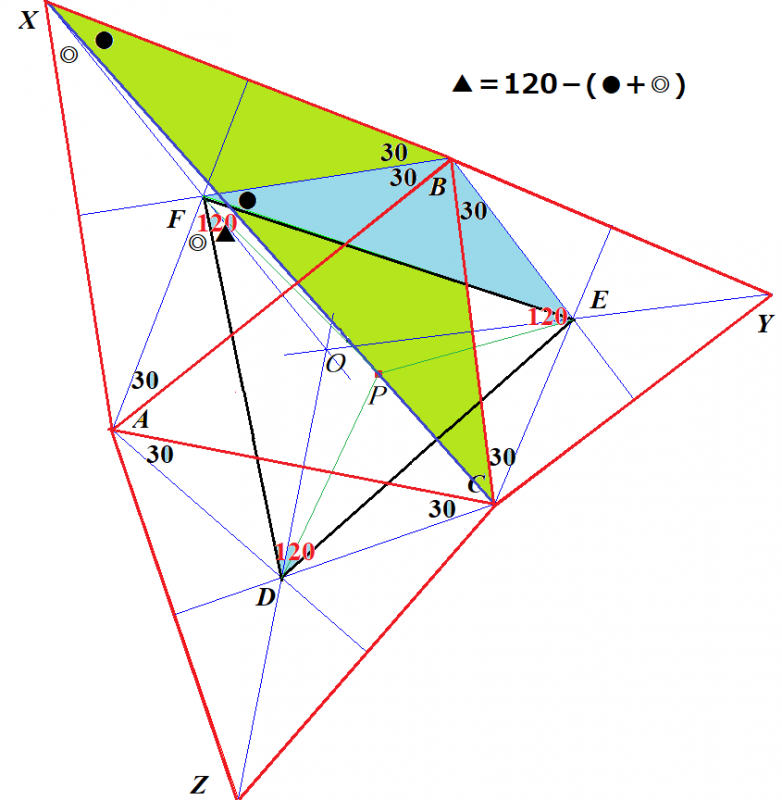
正3角形であるための必要十分条件は「3つの内角すべて(すくなくとも2つの内角)60°」です.
図形の対称性から,内角の1つが60°であることを証明すれば済みます.
これは,補助線1本引けば自明です.
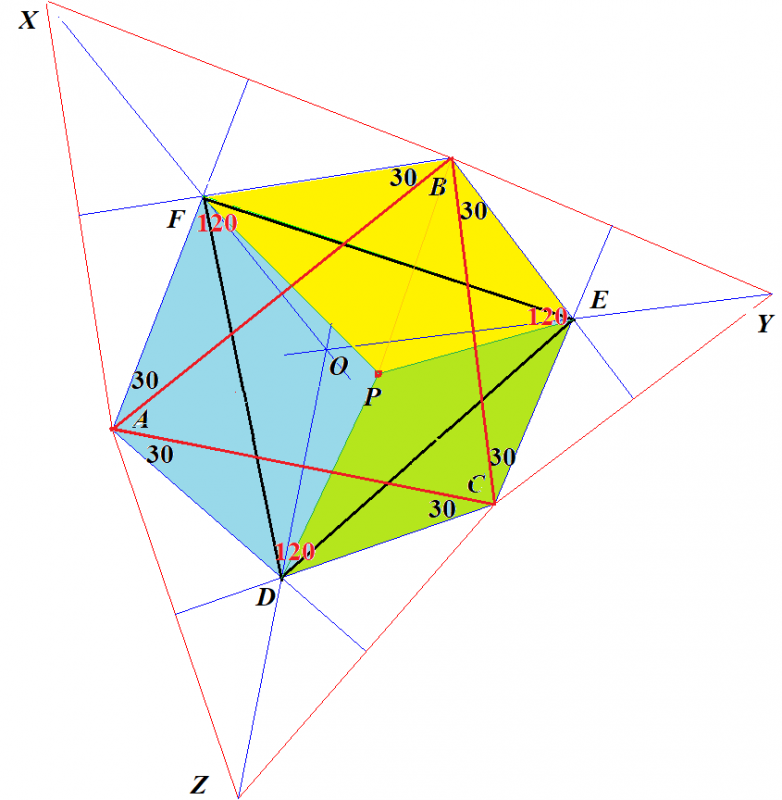
■さて,ここに出てきた6辺形には面白い性質があるのを見つけました.
この6辺形の中にある正3角形の中に点Pを中心になるように,正3角形の外の6辺形の部分を折り込むことができます.
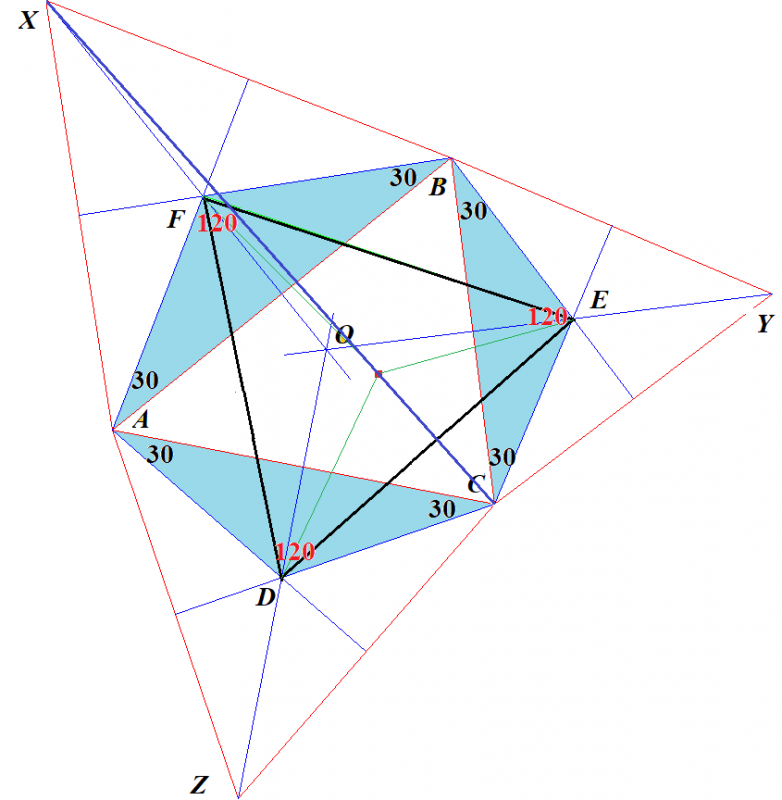
ナポレオンが発見したといわれるナポレオンの定理とは次のようなものです.
ナポレオンの定理
任意の⊿ABCの各辺上に正3角形を作図し,それら3つの正3角形の重心をD,E,Fとする.D,E,Fを結んでできる⊿DEFは正3角形である.
なかなか美しい形の定理ではありませんか,ナポオンの名を冠するのにふさわしい定理だと思います.ただし,ナポレオンが発見したかどうかは記録がなくわかりません.
ナポレオン (1769 -1821) は数学好きです.
陸軍幼年学校で,代数,三角法,幾何などを勉強し,数学で抜群の成績をおさめ,1784年にパリの陸軍士官学校に入学.数学が役に立つ砲兵科へと進みます.騎兵科,歩兵科でなく砲兵科に進んだのも戦術の時代の流れを見据えての決断でしょう.砲兵司令官,将軍,皇帝になりました.この時代にフランスには多くの数学者がいました.ラプラス,モンジュ,フーリエなどが近くにおり,エジプト遠征 (1798 年) にはモンジュやフーリエが同行しました.
ナポレオンは数学が大好き,このような幾何問題を考えるのが楽しみで,きっと定理を発見したのだと私は想像します.
証明
与えられた任意の3角形を⊿ABCとします.その各辺上に作図した正3角形のそれぞれの重心がD,E,Fです.
点Oは⊿ABCの垂心(⊿ABCのそれぞれの辺の垂直2等分線が交差する点)で,点D,E,Fは,この垂直2等分線上にあります.
色々な角の角度は図中に記入してあります.我々が証明すべきことは,⊿DEFが正3角形であることで,例えば,∠DFE=60を証明すれば済みます.考えてみてください.
ヒントは,∠BFE=∠BXC と ∠AFD=∠AXC を証明することです.
➡ 解答(証明)
反応拡散方程式
エンゼルフィッシュの縞模様やヒトデの星型はどうしてできるのでしょうか?
コンピュータの発明や暗号解読で有名な天才数学者アラン・チューリングが,”The chemical basis of morphogenesis”という論文を1952年に発表しました.今日,受精卵が細胞分裂を繰り返し分化し生物組織が出来ていく胚発生過程は遺伝子情報にプログラムされていることは公知です.1952年にチューリングが発表した理論は,「反応拡散系」が条件を満たせば,パターンや構造を自己成長形成するというものです.反応拡散系と言うのは,2つの物質(モルフォゲンと呼ぶ)が,反応し合いながら組織を介して拡散するもので,初期状態は均一であったものが,ランダムな外乱により,物質の濃淡の波が生じその波が生物の形や模様をつくりだすというものです.この数式でつくり出される模様は「チューリング・パターン」と呼ばれますが,コンピュータ・シミュレーションで描き出すと,条件により,動物の模様にそっくりな縞模様が出現したり,ヒトデの形を作ったりします.手の指が形づくられていくのは,その設計図が遺伝子により決定されているからと考えられていますが,もしかしたら,「指の形成はチューリングの理論のように波がつくっているのではないか」という論文が最近発表されたそうです.遺伝子はからだの構造の基本を決める設計図で,例えば,肺の形成の初期に気管支の分岐などを作るが,細かい肺胞の形成まではその設計図には書かれておらず,チューリング理論のように,現場の細胞同士のやり取り(反応と拡散)で作り上げられて行くのだろうと,近藤滋氏は言っています.
1952年に提唱されたチューリング理論は,現実の生物分野でそのような実験的証拠がなかったので,その後長い間,机上の空論と思われていました.1995年,近藤滋は,海洋エンゼルフィッシュのポマカンサスには,縞模様が皮膚に固定されていないことを発見しました.体の成長とともに,単純に比例して拡大する哺乳類の皮膚のパターンとは異なり,ポマカンサスの縞模様は,体の成長にともなうパターンの連続的な再配置が起こる.そして,縞間のスペースが維持されるという実験事実を観測しました.
実際,チューリング理論に基づくシミュレーションは,成長とともに形成されるパターンを正しく予測できたので,この理論の正しさを支持するものです.
■ チューリングの反応拡散系方程式
存在する2つの物質(モルフォゲン)が,反応したり拡散したりするのは,遺伝子情報で制御されるわけでもなく単純な化学反応で,以下の連立方程式で記述できます.u(t,r),v(t,r)は振動し,いろいろな形が形成されます.
■数学的補足
チューリングの反応拡散方程式の解の安定性を調べる数学について
(解が不安定(暴走)では,縞模様ができません)
しかしながら,テキストのメルマガで数式を多用することができませんので,
ここでは,言葉で説明するにとどめます.
微分方程式の解は指数関数であること.
反応項f,gはそれぞれ物質の濃度u,vの関数で,平衡点の周りでテーラー展開(1次の項まで)して線形化します.
このような連立線形微分方程式の性質は,ヤコビアンと呼ばれる行列Aで決まるが,
この行列Aの固有値の実部がすべて負であれば,解は安定になります.
行列Aの固有値を求めるのは面倒なので,条件を緩くして,行列Aの対角要素の和(固有値の和に同じ)が
負であるとして,さらに,拡散係数も0の場合から始めると,結局,f_u+g_v<0が得られます.
これは,促進剤と阻害剤が拮抗して働き,若干,阻害剤が強い条件を意味し,このようなときに縞模様が形成されます.
⇒ 数学追補
■Turing AM. The chemical basis of morphogenesis. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 237(641):37-72, 1952.
形態形成の主要な現象が,モルフォゲンと呼ばれる(互いに反応し組織を介して拡散する化学物質)の系で説明できることを示しました.初期状態は全く均一の系ですが,ランダムな外乱によって引き起こされる均一平衡の不安定性により,パターンや構造が発展する可能性があります.このような反応拡散系は,生物学的には珍しいシステムですが,数学的には便利な孤立した細胞のリングの場合にある程度詳細に考慮されます。
調査は主に不安定性の開始に関係しています。これが取ることができる6つの本質的に異なる形式があることがわかります。最も興味深い形では、定常波がリングに現れます。これは、たとえば、ヒドラの触手パターンや渦巻きの葉の原因となる可能性があります。球体上の反応と拡散のシステムも考慮されます。
そのようなシステムは原腸形成を説明しているようです。 2次元の別の反応システムは、垂れ下がった模様を思い起こさせます。また、2次元の定常波が葉序の現象を説明できることも示唆されています。
この論文の目的は、受精卵の遺伝子が,生物の解剖学的構造を決定する可能性のあるメカニズムを議論することです.この理論は新しい仮説を立てません.特定のよく知られた物理法則だけで多くの事実を説明するのに十分です.この論文を完全に理解するには、数学の十分な知識が必要です.若干の生物学,基礎化学,読者がこれらすべての専門家になることは期待できないので,多くの基本的な事実を説明しました(それらは教科書で見ることができますが,省略すると論文が読み難くなるので).
■Kondo S and Asai R. A reaction-diffusion wave on the skin of the marine angelfish Pomacanthus.Nature, 376(6543):765, 1995.
1952年にチューリングは,反応拡散系と呼ばれる分子的機構仮説を提案しました.これによると,均一な初期状態から周期的なパターンを作り出すことができます.形態形成でのパターン形成現象を説明するために,この反応拡散に基づいて多くの理論モデルが提案されましたが,生物学分野で,このような系の存在の決定的な実験的証拠はありません.海洋エンゼルフィッシュのポマカンサスには,皮膚に固定されていない縞模様があり,体の成長で単純に比例して拡大する哺乳類の皮膚のパターンとは異なり,ポマカンサスの縞模様はパターンの連続的な再配置により線間のスペースを維持します.パターンの変更はストライプの構造によって異なりますが,チューリング系に基づくシミュレーションプログラムは,将来のパターンを正しく予測できます.実際のパターンの再配置とシミュレートされたパターンの再配置の顕著な類似性は,反応拡散波がポマカンサスの縞模様の実行可能なメカニズムであることを強く示唆しています.
■Kondo S and Miura T. Reaction-diffusion model as a framework for understanding biological pattern formation.Science, 329(5999):1616-1620, 2010.
チューリング,または反応拡散(RD)モデルは,発達中の動物胚における自己制御パターン形成を説明する最も有名な理論モデルの1つです.実世界での関連は長い間議論されてきましたが,多くの説得力のある例がモデルを取り巻く懐疑論の多くを説得しつつあります. RDモデルはさまざまな空間パターンを生成できます.数学的研究により,それぞれに必要な相互作用の種類が明らかになり,さまざまな形態現象の実験での仮説として,このモデルが適用できる可能性があります.
このレビューでは,RDモデルが効果的に組み込まれている実験研究の例を使用して,モデルに不慣れな実験生物学者のためにこの理論の本質を説明します.